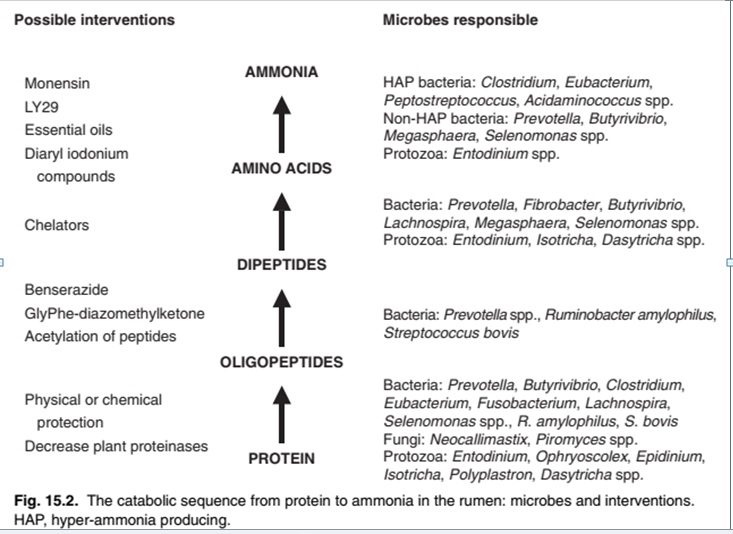
فرموله کردن احتیاجات اسیدآمینه در جیره غیرنشخوارکنندگان ساده تر از جیره نشخوارکنندگان است. زیرا نیازهای اسیدآمینه حیوانات غیر نشخوارکننده تنها از طریق پروتئین جیره تامین می شود. در مقابل نیازهای اسیدآمینه نشخوارکنندگان به وسیله مخلوط پروتئین جیره و پروتئین میکروبی حاصل می شود. بنابران بررسی متابولیسم اسیدآمینه در شکمبه به دلیل مفاهیم تغذیهای آن ضروری به نظر می¬رسد اسیدهای آمینه پس از آزاد شدن در شکمبه، به مدت طولانی به فرم آزاد باقی نمیمانند. آنها یا به پروتئین میکروبی تبدیلشده و یا به آمونیاک تجزیه میشوند. برای به حداکثر رساندن بازدهی تولید اسیدآمینه از شکمبه، باید سرعت تجزیه پروتئین خوراک کم باشد تا جاییکه تمام پروتئینی که تخریب میشود بهوسیله میکروبها استفاده شود. تجزیه پروتئین پروتئین منبع فراوانی از نیتروژن در اکثر جیرههای نشخوارکنندگان است. پروتئین غذا بهسرعت توسط پروتئازهای میکروبی ترشح شده توسط باکتریها، پروتوزوا و قارچهای موجود در شکمبه، بهصورت پپتیدها و اسیدهای آمینه در آمده و سرانجام توسط سلولهای میکروبی جذب میشوند (کوتا و راسل، ۱۹۹۷). و درنهایت تعدادی از عوامل که بر میزان دقیق و مقدار تجزیه تأثیرگذارند، ارزش غذایی پروتئین را تعیین میکنند (والاس و همکاران، ۱۹۹۷). ویژگیهای شیمیایی و فیزیکی مختلف پروتئین، مانند حلالیت، نوع ساختار دوم و سوم و حضور پیوندهای دی سولفیدی (والاس، ۱۹۸۳) عوامل تعیینکننده مهم قابلیت هضم پروتئین توسط میکروارگانیسمهای شکمبه هستند. ازآنجاییکه برخی از پروتئینها سریعتر از بقیه در شکمبه تخمیر میشوند (والاس، ۱۹۹۴)، نرخ و میزان تجزیه پروتئین برای تولید پپتیدها و اسیدهای آمینه که سپس به پروتئینهای میکروبی تبدیلشده یا توسط میکروارگانیسمهای شكمبه به آمونیاك تجزیه میشوند، ترکیب آمونیاک در شکمبه را بهطور قابل توجهی تحت تأثیر قرار میدهند (هریستو و برودریک، ۱۹۹۴). هریستوف و برودریک (۱۹۹۴) دریافتند که پروتئینهای محلول مانند کازئین، اسید آمینه بیشتر و آمونیاک کمتری را برای تولید پروتئینهای میکروبی ارائه میدهند. تعدادی از انواع مختلف آنزیمهای پروتئولیتیک تولید و ترشح شده توسط بسیاری از گونههای پروتوزایی مژکدار، باکتریها و قارچهای پروتئولیتیک شکمبه یافت شده اند (والاس، ۱۹۹۴؛ والاس و همکاران، ۱۹۹۷). همچنین نوع جیره تأثیر عمدهای بر فعالیت پروتئولیتیک و گونههای میکروبی مسئول آن فعالیت دارد (ناگنت و منگان، ۱۹۸۱؛ سیدونس و پارادیاین، ۱۹۸۱؛ هازلوود و همکاران، ۱۹۸۳). گزارش شده است که گیاهان تازه سبب افزایش فعالیت پروتئولیتیک بیشتر نسبت به جیرههای خشک میشوند (ناگنت و منگان، ۱۹۸۱؛ هازلوود و همکاران، ۱۹۸۳؛ ناگنت و همکاران، ۱۹۸۳). همچنین بیان شده است جیرههای حاوی غلات نسبت به جیرههای خشک، فعالیت پروتئولیتیک بالاتری را ایجاد میکنند، احتمالاً به این دلیل که این جیرهها بیشتر باعث تکثیر گونههای آمیلولیتیک شکمبه می¬شوند که فعالیت پروتئولیتیک بیشتری نسبت به سلولولیتیک¬ها دارند (سیدونس و پارادیاین، ۱۹۸۱). قابل توجه است که مدل آنزیمهای پروتئولیتیک در حیوانات با یک جیره یکسان و یا مشابه، با یکدیگر متفاوت است (والاس و همکاران، ۱۹۹۷؛ فالکونر و والانس، ۱۹۹۸). پیشنهادی مطرح شده است که پروتئولیز نه تنها توسط میکروارگانیسمهای شکمبه، بلکه توسط پروتئازهای اندوژنوس گیاهی نیز انجام میشود که ممکن است نقش مهمی در تجزیه پروتئینهای گیاهی تازه در شکمبه ایفا کند (ژو و همکاران، ۱۹۹۹ ) پروتئینازهای گیاهی نقش مهمی در تجزیه پروتئین در سیلو ایفا میکنند و منطقی است که آنها ممکن است در شکمبه نشخوارکنندگان چراکننده نیز نقش داشته باشند. تجزیه پپتید پپتیدها مواد حد واسط در تبدیل پروتئین¬ها به آمونیاک هستند. میکروارگانیسم های شكمبه، پپتيدهاي مختلف را به میزان متفاوتی تجزيه میکنند (والاس و همكاران، ۱۹۹۳). هنگامیکه پروتئینها سریع تجزیه شوند، پپتیدها در شکمبه انباشته میشوند، ۲-۱ ساعت پس از تغذیه به پیک غلظت رسیده و پسازآن کاهش مییابند (برودریک و والاس، ۱۹۸۸). چن و همکاران (b، ۱۹۸۷) گزارش دادند که باکتریهای شكمبه، پپتیدهای آبدوست را نسبت به پپتیدهای آبگریز، سریعتر تجزیه میکنند. هیدرولیز پپتیدها در شکمبه شامل دو مرحله می باشد. اولين مرحله فعاليت آمینوپتیدازی است كه بهجای جدا شدن اسیدهای آمینه به صورت تکی، زنجيره پپتيدی به دي پپتيدها شکسته میشود (والاس، ۱۹۹۶). در مرحله بعد دی پپتیدها به آمینواسیدها تقسیم میشوند.
تجزیه آمینو اسید تجزیه اسید آمینه مرحله نهایی تبدیل پروتئین جیره به آمونیاک است. میزان تخریب اسید آمینه معمولا بیشتر از مصرف اسید آمینه توسط میکروارگانیسم های شکمبه است، اسید های آمینه اضافی مهم ترین منبع آمونیاک در شکمبه هستند غلظت اسیدهای آمینه در مایع شکمبه حتی پس از تغذیه نیز کم می¬باشد. این مشاهدات ممکن است گمراه کننده باشند، زیرا میزان اسیدهای آمینه باند شده در پپتید که توسط سلول های میکروبی گرفته شده اند ممکن است بسیار متفاوت از اسیدهای آمینه آزاد مربوطه باشند. غلظت اسیدهای آمینه در شکمبه به ماهیت جیره بستگی دارد. بیشترین میزان پس از تغذیه یونجه خشک مشاهده شده است. برودریک و والاس (۱۹۸۸) انباشتگی اسیدهای آمینه هنگام تجزیه سریع پروتئین (نه تخریب آرام) رخ می دهد. هینو و راسل (۱۹۸۵) فعالیت دآمینازی میکروارگانیسم های سالم و مایعات سلولی را مقایسه و نتیجه گرفتند که ظرفیت بالای دآمیناسیون نشان می دهد که میزان جذب پپتیدها یا اسیدهای آمینه به سلول ها ممکن است میزان تولید آمونیاک را محدود کند. منبع: Amino Acids in Animal Nutrition, Second Edition (۲۰۰۳), Edited by J.P.F. D’Mello, Formerly of the Scottish Agricultural College Edinburgh, UK